![]()
![]()

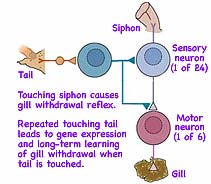
 Starting shortly after 1960, Eric Kandel (b1929 in Vienna, Austria, driven out of Europe in 1939 by Nazi Germany, shown at right at Columbia U→) began studying the foot-long sea snail, Aplysia californica (←at left). This is a simple mollusk with only a few, but giant, neurons which allow investigation of changes in a single neuron caused by learning. While many doubted it at that time, there is much in common with learning changes which occur in our brains. (For his discoveries Kandel won 1/3 of the 2000 Nobel Prize in Physiology/Medicine.) The snail instinctively retracts its delicate (darker) gill tissues when endangered. Just as humans do when subjected to a repeated stimuli, the snail learns to react less and less to repeated light touches to its syphon. This retraction response involves only 24 sensory neurons and 6 connected motor neurons. The neurons and their interconnecting neural circuitry are the same in every sea snail. (So how can learning occur in a neural circuit which is precisely wired?)
Starting shortly after 1960, Eric Kandel (b1929 in Vienna, Austria, driven out of Europe in 1939 by Nazi Germany, shown at right at Columbia U→) began studying the foot-long sea snail, Aplysia californica (←at left). This is a simple mollusk with only a few, but giant, neurons which allow investigation of changes in a single neuron caused by learning. While many doubted it at that time, there is much in common with learning changes which occur in our brains. (For his discoveries Kandel won 1/3 of the 2000 Nobel Prize in Physiology/Medicine.) The snail instinctively retracts its delicate (darker) gill tissues when endangered. Just as humans do when subjected to a repeated stimuli, the snail learns to react less and less to repeated light touches to its syphon. This retraction response involves only 24 sensory neurons and 6 connected motor neurons. The neurons and their interconnecting neural circuitry are the same in every sea snail. (So how can learning occur in a neural circuit which is precisely wired?)
Kandel found that the snail can also associate a stimulus to its tail applied simultaneous with a light touch to its gills or siphon. By associating the simultaneous stimuli, the snail learns to withdraw its gills in response to just the stimulus to the tail. When its tail is stimulated, the neurotransmitter serotonin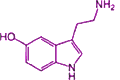 (also found in human brains; its structural formula shown at right→) is released at the terminal synapse of the sensory neuron carrying message from the tail. This activates receptors which release cyclicAMP which function as a secondary messengers. The cyclicAMP activates protein kinases which, by adding phosphate groups, prepare more neurotransmitter for release, boosting the capacity to carry the signal across the synapse to the motor neuron. No new proteins are synthesized, but for a short time the tail will have heightened sensitivity. If the stimulus is more forceful the reflex is stronger. This amplification lasts only for a few minutes, demonstrating short-term memory.
(also found in human brains; its structural formula shown at right→) is released at the terminal synapse of the sensory neuron carrying message from the tail. This activates receptors which release cyclicAMP which function as a secondary messengers. The cyclicAMP activates protein kinases which, by adding phosphate groups, prepare more neurotransmitter for release, boosting the capacity to carry the signal across the synapse to the motor neuron. No new proteins are synthesized, but for a short time the tail will have heightened sensitivity. If the stimulus is more forceful the reflex is stronger. This amplification lasts only for a few minutes, demonstrating short-term memory.
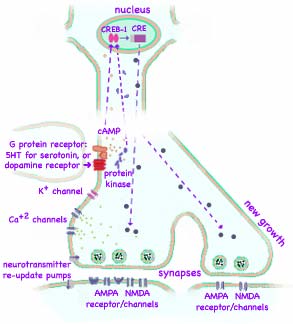
 But if repeatedly stimulated, long-term memory is created. Converting short-term memory to long-term memory requires appropriately spaced repetition of the stimulus. The additional serotonin caused by repeated stimulation also results in higher concentration of the second messenger (cyclicAMP) which activates additional protein kinases. These along with another enzyme migrate into the nucleus of the neuron. There they activate genes in the sensory neuron resulting in production of new protein used to construct additional synaptic connections. (Kandel found that a cAMP-response-element-binding protein, CREB-1 for short, turns on the genes.) When the forceful stimulus is repeated several times, the sensitization may remain for weeks. What we call memory is actually additions to the billion of synapses that form the contact points between the neurons. While genes are involved in the growth of new synaptic connections, it is the stimulation by the environment that governs the expression of the genes and causes the learning. Kandel was able to actually count new synaptic connections in both sensory and motor neurons after learning. So in a real sense, the connections between neurons in our brains actually grow and change when we have experiences and learn. This is not a new idea. Santiago Ramón y Cajal proposed to the Royal Society in 1894 that memory might be stored in the growth of new connections. But lots of more complex ideas had been proposed in the meantime. Now there is evidence that his proposal matches reality.
But if repeatedly stimulated, long-term memory is created. Converting short-term memory to long-term memory requires appropriately spaced repetition of the stimulus. The additional serotonin caused by repeated stimulation also results in higher concentration of the second messenger (cyclicAMP) which activates additional protein kinases. These along with another enzyme migrate into the nucleus of the neuron. There they activate genes in the sensory neuron resulting in production of new protein used to construct additional synaptic connections. (Kandel found that a cAMP-response-element-binding protein, CREB-1 for short, turns on the genes.) When the forceful stimulus is repeated several times, the sensitization may remain for weeks. What we call memory is actually additions to the billion of synapses that form the contact points between the neurons. While genes are involved in the growth of new synaptic connections, it is the stimulation by the environment that governs the expression of the genes and causes the learning. Kandel was able to actually count new synaptic connections in both sensory and motor neurons after learning. So in a real sense, the connections between neurons in our brains actually grow and change when we have experiences and learn. This is not a new idea. Santiago Ramón y Cajal proposed to the Royal Society in 1894 that memory might be stored in the growth of new connections. But lots of more complex ideas had been proposed in the meantime. Now there is evidence that his proposal matches reality.
 (Two relevant questions of memory are (1) Where is memory stored? and (2) How is it stored?) Explicit memory which requires conscious attention to people, objects and places, facts and events, is stored in the medial temporal lobe of the hippocampus in the brain. Implicit, procedural memory which does not require conscious attention such as skills, habits, and non-associative and associate learning resides in the striatium, amygdala, cerebellum, and reflex pathways. Both kinds of memory start as short-term lasting typically minutes, but by repetition can be converted to long term memory lasting at least days or weeks. Only long-term memory involves synthesis of new protein.
(Two relevant questions of memory are (1) Where is memory stored? and (2) How is it stored?) Explicit memory which requires conscious attention to people, objects and places, facts and events, is stored in the medial temporal lobe of the hippocampus in the brain. Implicit, procedural memory which does not require conscious attention such as skills, habits, and non-associative and associate learning resides in the striatium, amygdala, cerebellum, and reflex pathways. Both kinds of memory start as short-term lasting typically minutes, but by repetition can be converted to long term memory lasting at least days or weeks. Only long-term memory involves synthesis of new protein.
There is for each of our senses, for touch, for vision, for smells, for hearing a topographical map-like re-creation in the brain, an internal representation that is organized so that relationships are preserved. For example, one can ask an experienced person how to get from one known location to another, or to recall an event from the past and they will be able to recreate from their memories images of the necessary route, or images from the event. These relationships are stored as synapse connections in the brain. As a result of memories, those with more experiences will have a larger right hippocampus than those inexperienced! Other animals such as mice form particularly good spacial maps for remembering their surroundings and experiences. Kandel's team have used this to investigate differences between the explicit memory of mice and the implicit memory of the sea snail. ![]() While dopamine (structural formula at right→) is the neurotransmitter instead of serotonin, the chemistry is nearly identical. And while the unconscious long-term memory requires repeated stimulation from the environment, long-term explicit memory of a spacial map requires conscious attention from the prefrontal and posterior parietal cortex. Apparently if you don't have the motivation and are not willing to pay attention, you aren't going to remember very long! The equivalent of the repetition needed to learn skills is motivation and attention for explicit learning. But while attention is expanding some memory, it is likely also depressing other memories which we judge more trivial.
While dopamine (structural formula at right→) is the neurotransmitter instead of serotonin, the chemistry is nearly identical. And while the unconscious long-term memory requires repeated stimulation from the environment, long-term explicit memory of a spacial map requires conscious attention from the prefrontal and posterior parietal cortex. Apparently if you don't have the motivation and are not willing to pay attention, you aren't going to remember very long! The equivalent of the repetition needed to learn skills is motivation and attention for explicit learning. But while attention is expanding some memory, it is likely also depressing other memories which we judge more trivial.
(Is the memory thus stored in the entire neuron cell or a more limited location?) CREB functions as a molecular switch that turns short-term memory to long-term memory by generating proteins and messenger RNA which spread throughout the neuron. However, Kandel found that many mRNA remain dormant until repeat synapse use arouses the mRNA to build synapse building proteins. Team member Kelsey Martin found that while a neuron might have thousands of synapse, only those synapses activated by recent use can utilize the new protein to grow new connections. She found that not only is the cyclicAMP-dependent protein kinase released by the dopamine neurotransmitter necessary to develop new connections, but a local protein assembly mechanism governed by about 250 messenger RNA must also be available. If local protein assembly is not functioning, the synapse growth initially occurs, but rapidly regresses.
Another team member, Kausik Si, a postdoc in Kandel's laboratory, found a protein called CPEB (Cytoplasmic Polyadenylation Element Binding protein) which functions as a selective trigger which activates dormant mRNA at active synapses. Generally, such protein has attached phosphate groups which activate mRNA. However the CPEB in the neurons of snails, flies and mammals lack those phosphates. Instead, it has a strange functional domain unusually rich in the amino acid glutamine and lacks any secondary structure. Only one type of protein has a similar domain: prions [proteinaceous infectious particle: an unusual type of protein which can shape-shift into insoluble clumps and catalyze other prions to follow suit. These clumps are resistant to normal methods of denaturing proteins such as heat, radiation, formaldehyde, and protease enzymes. Prions cause brain and neuron diseases such as scrapie (in sheep), chronic wasting disease, (in deer and elk), Creutzfeldt-Jakob disease (CJD) and Kuru (both in humans), and bovine spongiform encephalopathy (mad cow disease) ]. The self-perpetuating property of pions might, by anchoring to a site and working cooperatively to sustain a memory at a synapse, keep a memory vivid over time. After consulted with prion expert Susan L. Lindquist, Kandel, Lindquist, and Si proposed a model of memory storage in the December 26, 2003, issue of Cell, accompanied by a Kandel and Si article on the role of CPEB long-term memory storage in Aplysia. Lindquist and Kandel speculate further that a similar prion mechanism might be involved in developmental processes and cancers, where cells also maintain a continuing function. Si has confirmed the CPEB in the snail neurons does act like a prion. While very active in the prion clusters, it barely functions in the nonprion state. Perhaps certain events such as repeating the times tables, practicing piano, and one's first romantic experiences become so unforgettable by triggering enough CPEB production so that some convert into prions and perpetuate themselves at the synapses where they prevent future use of that synapse from destroying or modifying the long-term memory forms.
In 2002 Todd Sacktor identified another protein kinase (protein kinase M zeta, or PKMζ for short) required to maintain long term memory. Its role was verified by injecting into mice a small synthetic protein which blocked PKMζ. The result was that the mice forgot what they had previously learned. Once the synthetic protein was gone from the mice and the PKMζ was again functioning, the mice were able to re-learn and remember. Such a molecule as this synthetic protein could have value for situations where it would be desirable to forget a bad experience or habit.
While we are beginning to understand how long-term memory is formed, stored and perpetuated, there remains much to be learned about the interactions and other processes within the brain.
There are many techniques for focusing one's attention to assist is forming long-term memory. Perhaps it might be valuable at this time to compile a description of many of these techniques and distinguish their characteristics and more valuable features. For example, outlining the main mechanisms, constructing Venn diagrams and making lists of new vocabulary for this and earlier pages about the nervous system could all be useful but each highlights different aspects.
One of the more crucial memory functions is pattern recognition: A great part of the success of animal life, particularly that of humans, is based on recognizing patterns. It is crucial that animals recognize what is suitable food, and what situations constitute danger to their survival. We recognize where we are, and what property is ours. We recognize what tool can help us with a needed task, and we recognize when a weather change suggests an appropriate response from us. When Altzheimers disease ravages human minds and people no longer recognize even the faces of family, they require much care and supervision just to keep living.
Facial recognition emphasizes several crucial characteristics of pattern recognition. Every face changes. We may smile or frown, parts may sag as we tire, our hair changes length, style, and sometimes color, and our nose grows and skin wrinkles as we age. Yet we often recognize and distinguish one face from another despite these changes. So pattern recognition is not a matter of identifying things which are identical, but of deciphering which are significant characteristics which need to be similar, and which are insignificant and should be ignored.
Still, not all pattern recognition is useful. For example it was recently noted that plasma in the space between stars forms patterns like the alpha helix of DNA. (This is because charged particles crossing magnetic fields such as in the space between stars have a force exerted upon them perpendicular to their motion. The helix of DNA is due to bond angles and spacing of atoms in the molecule.) Because the two phenomena have entirely different causes, there is little value in recognizing their similar pattern. Perhaps there is generally little value in noting similar patterns in phenomena at vastly different scales and situations. As with many generalities, there have been exceptions: After finding recognizable animal patterns in the constellations, earlier humans learned they could be used to tell the seasons, navigate across oceans without recognizable landmarks, and learn more about the heavens. Those would have been much more difficult without recognizing the patterns even though patterns such as Ursa Major and Minor are not remotely related to real bears.
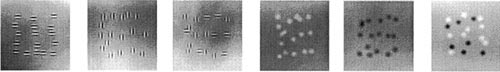 Our nervous systems can recognize patterns such as the letter
Our nervous systems can recognize patterns such as the letter E
when it is rotated, composed incompletely of various components, involves components out of place, or of various composition. Parameters such as component separation, continuity, and pattern fit play an important role in recognizing and distinguishing a figure from its background. The foveal, the center of focused vision in our eyes, can perceive patterns close to the theoretical limits of the position and number of features needed to define the pattern; however peripheral vision is considerably less capable, requiring many samples to reliably determine a pattern. Laurent Itti, Christof Koch and Jochen Braun have suggested that spatial vision thresholds reflect a single level of preprocessing which may occur in the eye or later in the visual cortex of the brain.
You might develop an investigation to further investigate recognition of such patterns.
The process of looking for something and then recognizing it is considerably more complex. That challenge requires recognizing and paying attention to the most relevant of visible features from around 108 bits of information being detected by our retina each second. Rarely do we, or even a robot Martian lander, open our eyes without a preconceived idea of what will be in view based on location and memories of previous images from similar situations. That preconception provides a bias of what patterns and features should appear and be relevant, so as to provide one or more targets to try to recognize. (For example, above it helped to know we were looking for the letter E.) Vidhya Navalpakkam and Laurent Itti have proposed a model where the most active feature visible is then selected for attention based on the preconceived expected scene. No intelligence is involved in this selection as the selection is based strictly on the preconceived expectation. The brain then attempts to compare the visual image at this location with memories of similar patterns to identify the object and any change. These parallel processed comparisons include evaluating the feature to prior visual memories, relevance to task at hand and importance. In the final stage the brain updates the expectation. For example, if the object is of low relevancy, it is marked trivial and will not be chosen as the active feature in the next iteration. The process rapidly selects task-relevant targets bases on a given task definition. Using such a visual search behavior, many targets are recognized amidst clutter and diverse backgrounds then classified into corresponding categories, generally with few errors. The visual system learns the task-relevant locations in an incremental manner, and then uses the learned target features as well as likely target locations to bias the attentional system to guide attention towards the target. Visual feature maps are computed in parallel. Adjusting the weights of the low-level feature maps promote the target's relevant features and suppress irrelevant features. Visual features are stored in visual working memory along with links to symbolic knowledge which deals with high-level concepts and objects.
Life is full of surprises: a great Christmas gift, a new trick, wardrobe malfunctions, reckless drivers, terrorist attacks, and tsunami waves. Survival requires that we rapidly identify surprising events and determine prudent courses of action. Yet we have little understanding, vocabulary, or quantitative unit for measuring surprise. Only unexpected stimuli from our senses are transmitted along nerves to the brain. The sensory cortex may have evolved to predict and to trivialize the expected regularities of the world, focusing our attention instead on events that are unpredicted and surprising. Evidence from visual, olfactory, and auditory cortices, and retinal ganglion cells and cochlear hair cells show that respective nerve signals greatly diminish with repeated or prolonged exposure to an initially novel stimulus of each of those senses. Surprise and novelty are central to learning and memory formation; surprise seems to be a necessary trigger for associative learning. Preliminary evidence suggests the hippocampus has a role as a novelty detector.
Communicating technical information such as observations and findings is a skill used by scientists but useful for most others. If you need course credit, use your observations in your journal to construct a formal report.
![]()
to next investigation
to Biochemistry menu
to e-Chemistry menu
to site menu