![]()
![]()
Light presents us with a paradox. The ancient Greeks proposed that that objects sprayed out particles of light in reaction to our gazes. Isaac Newton amplified the concept of light particles in an attempt to explain color effects. His contemporary, Thomas Young proposed that light flowing as waves could better explain the color effects. Later Albert Einstein found that bundles of light energy (called photons) were needed to explain the photoelectric effect. So we are left with the dualism paradox where light exhibits both wave and particle properties.
Different colors can be thought of as either waves with different separation (wavelength = λ) or different sized energy bundles (E = hν where ν is the frequence of passing waves with a constant, h). Either way, color comes in an infinite continuum of different colors. Many organisms do not distinguish different colors of light but just detect a limited range of colors. All vertibrates including humans have both more sensitive light detectors (called rods) that do not distinghish color (so we only see shades of gray at night), and additional, less sensitive detectors (called cones) which do distinguish color. Like all other forms of life, we only see a limited range of colors which we label red to violet. Rather than actually measuring either the light's actual color and intensity, the detectors in our eyes only distinguish the relative intensities of three bands of colors. (Some fish, frogs, birds, and turtles see as many as five distinct color bands. Other mammals see two color bands) Our nervous system attempts to interpret that very limited data to provide our perception of color. This mechanism which we chauvinistically call primary colors is often inaccurate! But it allows for inexpensive color printing and video presentations using as few as three inks or phosphors. While it is sometimes erroneously said that cyan, yellow, and magenta pigments or red, green and blue lights can create full color images, it would be more accurate to say those provide an illusion to us of providing all the colors in the full visible spectrum.
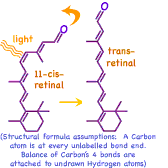
As long suspected, the orange pigment common in carrots and other fruits and vegetables, β-carotene, is a key ingredient in vision. The symmetrical β-carotene molecule is split in half and the new ends slightly oxidized forming two identical molecules of an alcohol called Vitamin A. Each new end is further oxidized in the eye to an aldehyde (←diagramed at left). It is this 11-cis-retinal molecule which actually absorbs the light, triggering the nerve process we call vision. 11-cis-retinal has double bonds alternating along the tail-like chain which makes the molecule stiff. When the 11th bond absorbs light's energy, the double bond's second electron pair dissociates allowing the remaining single bond to momentarily freely rotate. Roughly 50% of the time the double bond reforms in the cis configuration without any further effect. But the double bond can also reform in the trans configuration matching the neighboring double bonds. This straightens the kink in the molecule's shape, leaving an essentially straight tail attached to the ring.
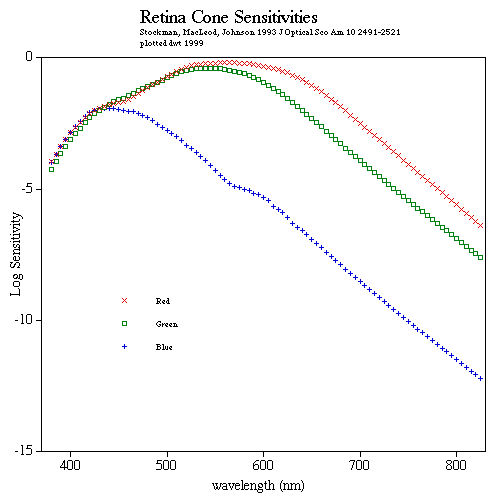
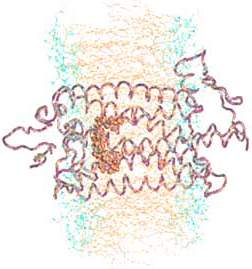 This shape transformation causes the surrounding protein complex, called opsin (depicted at right→), to initiate the nerve impulse. Opsin is a mostly coiled, seemingly tangled protein, a member of a group, all of which thread across a double lipid membrane exactly 7 times. (Here the lipid's middle layer of long non-polar hydrocarbon chains are colored tan and the polar functional groups which form the inner and outer surfaces are shown blue.) These 7-transmembrane proteins are part of a still larger family called G-proteins (which get their name because all of them bond to the nucleotide GMP, guanosine monophosphate). Some (such as those involved in detecting taste and odors) are triggered by binding to another ligand. In this case the ligand is 11-cis-retinal which stabilizes the opsin and keeps it inactive. The shape change of the retinal causes a cascade of molecular changes (similar to that diagrammed in Biochemistry 3) which activate a partner G-protein, transducin, which serves to magnify the signal. The complex is regenerated by releasing the trans-retinal and binding to a new 11-cis-retinal. The shape change of the retinal is among the fastest known photochemical reactions taking 200 fs (1 fs = 10-15 sec). The complete cycle requires milliseconds until the complex is ready to detect the next photon of light. The 11-cis-retinal is identical in all rods and cones. Nerve responses carrying signals from cones shows that there are three different sensitivity curves for various colors of light. So the difference in sensitivities to different colors of light, which makes color vision possible, is due to a few differences in amino acids in key portions of the opsin which modify the sensitivity of the encased 11-cis-retinal. (Note 11-cis-retinal inside opsin above.)
This shape transformation causes the surrounding protein complex, called opsin (depicted at right→), to initiate the nerve impulse. Opsin is a mostly coiled, seemingly tangled protein, a member of a group, all of which thread across a double lipid membrane exactly 7 times. (Here the lipid's middle layer of long non-polar hydrocarbon chains are colored tan and the polar functional groups which form the inner and outer surfaces are shown blue.) These 7-transmembrane proteins are part of a still larger family called G-proteins (which get their name because all of them bond to the nucleotide GMP, guanosine monophosphate). Some (such as those involved in detecting taste and odors) are triggered by binding to another ligand. In this case the ligand is 11-cis-retinal which stabilizes the opsin and keeps it inactive. The shape change of the retinal causes a cascade of molecular changes (similar to that diagrammed in Biochemistry 3) which activate a partner G-protein, transducin, which serves to magnify the signal. The complex is regenerated by releasing the trans-retinal and binding to a new 11-cis-retinal. The shape change of the retinal is among the fastest known photochemical reactions taking 200 fs (1 fs = 10-15 sec). The complete cycle requires milliseconds until the complex is ready to detect the next photon of light. The 11-cis-retinal is identical in all rods and cones. Nerve responses carrying signals from cones shows that there are three different sensitivity curves for various colors of light. So the difference in sensitivities to different colors of light, which makes color vision possible, is due to a few differences in amino acids in key portions of the opsin which modify the sensitivity of the encased 11-cis-retinal. (Note 11-cis-retinal inside opsin above.)
The strong similarities among molecules which detect aspects of the environment (such as light, taste, odor, and pressure) and convert that information to nerve impulses may be apparent when earlier presentations are compared to the description above. This contrasts with the very different organs and their cellular structures than are involved.

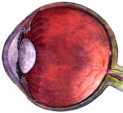 The amount of light enters the nearly spherical eye (diagrammed at right→) is controlled by an iris, a circular ring shaped shutter which opens and closes depending on light intensity. Once the light enters the eye through the iris, little is allowed to escape back out. This makes the variable-sized transparent center of the iris at the front of the eyeball to appear black. The iris itself contains a variety of pigments to block excess light from entering the eye. The mix of pigments give the ring of the iris its unique, genetically determined dark color (blue eyed, brown eyed, etc.).
The amount of light enters the nearly spherical eye (diagrammed at right→) is controlled by an iris, a circular ring shaped shutter which opens and closes depending on light intensity. Once the light enters the eye through the iris, little is allowed to escape back out. This makes the variable-sized transparent center of the iris at the front of the eyeball to appear black. The iris itself contains a variety of pigments to block excess light from entering the eye. The mix of pigments give the ring of the iris its unique, genetically determined dark color (blue eyed, brown eyed, etc.).
In addition to the intensity of light being controlled by the iris, the cone photoreceptors can also adapt to the brightness of light. And signal processing neurons in the eye can further adjust for light intensity. Similarly the rods and their neural circuitry also adapt to diminished light intensity.
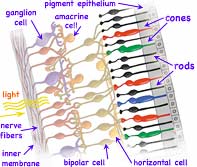 The light reaches the eye by traveling diverging, presumably straight-line paths outward from its origin. This assumption our brain later uses to determine both the source direction and distance. Upon entering the eye, a series of fluid and semi-rigid lenses (shown grey at left side of eyeball above) refocus the previously diverging light paths so those with identical origin arrive at a common spots on the retina on the far back of the eye (far right in diagram above). (←The retina is shown enlarged at left, and enlarged several more orders of magnitude below right ↓) The light passes through 0.5 mm of nearly transparent nerves and related structures (such as the nuclei, mitochondria and ribosomes of the light detecting neurons), finally reaching and being absorbed in layers of rod or cone shaped pigment. Here multiple folded membranes form discs containing the opsin pigment complex (also called visual purple due to its actual appearance. ←In the diagram at left, cones are falsely colored as a reminder of their three, slightly different color sensitivities.) Further towards the outside of the eye lies an additional pigmented epithelium layer which absorbs any remaining light that otherwise could scatter back blurring the image the eye is detecting. A Voltage varied signal is sent inwards from the light detecting rods and cones to a collection in primates of at least 22 types of ganglions and other signal processing neurons. These do initial image processing before sending signals along one of more than a million nerve fibers composing the large optic nerve to the back of the brain for further processing and perception.
The light reaches the eye by traveling diverging, presumably straight-line paths outward from its origin. This assumption our brain later uses to determine both the source direction and distance. Upon entering the eye, a series of fluid and semi-rigid lenses (shown grey at left side of eyeball above) refocus the previously diverging light paths so those with identical origin arrive at a common spots on the retina on the far back of the eye (far right in diagram above). (←The retina is shown enlarged at left, and enlarged several more orders of magnitude below right ↓) The light passes through 0.5 mm of nearly transparent nerves and related structures (such as the nuclei, mitochondria and ribosomes of the light detecting neurons), finally reaching and being absorbed in layers of rod or cone shaped pigment. Here multiple folded membranes form discs containing the opsin pigment complex (also called visual purple due to its actual appearance. ←In the diagram at left, cones are falsely colored as a reminder of their three, slightly different color sensitivities.) Further towards the outside of the eye lies an additional pigmented epithelium layer which absorbs any remaining light that otherwise could scatter back blurring the image the eye is detecting. A Voltage varied signal is sent inwards from the light detecting rods and cones to a collection in primates of at least 22 types of ganglions and other signal processing neurons. These do initial image processing before sending signals along one of more than a million nerve fibers composing the large optic nerve to the back of the brain for further processing and perception.
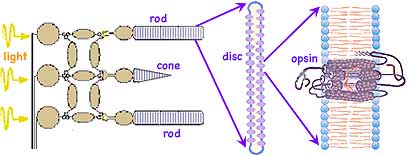 Each human retina contains about 108 rod cells and roughly 5 x 106 cone cells. The rods, which are most plentiful around the periphery of the retina, can detect and count single photons of light, sending a signal to neurons which integrate signals from neighboring rods. This provides for the detection of very faint light and the ability to notice movements at the edges of our vision. But that integration process creates as much as 1/10 second delay. The cones are highly concentrated on the back side of the eye, within a couple degrees of directly opposite from the iris and lenses in an area responsible for our primary visual image. This central area also has many more ganglion and other neurons to process the signals from the cones. (Just below this central area is a
Each human retina contains about 108 rod cells and roughly 5 x 106 cone cells. The rods, which are most plentiful around the periphery of the retina, can detect and count single photons of light, sending a signal to neurons which integrate signals from neighboring rods. This provides for the detection of very faint light and the ability to notice movements at the edges of our vision. But that integration process creates as much as 1/10 second delay. The cones are highly concentrated on the back side of the eye, within a couple degrees of directly opposite from the iris and lenses in an area responsible for our primary visual image. This central area also has many more ganglion and other neurons to process the signals from the cones. (Just below this central area is a blind spot
where the axons meeting to form the optic nerve bundle leading to the brain preclude rods and cones being located there.) While different in shape, both rods and cone cells produce flat disk shaped folded membranes rich in opsin, which fill the rods and cones. While the cells themselves are long-lived, they slough off the outer most disks as new ones are formed. The major portion of the 7-transmembrane opsin is believe to be manufactured using the Golgi apparatus in the transparent inner segments
of the neurons. The 11-cis-retinal enters from the neighboring pigmented epithelium and may be recycled there after each light induced structural transformation. (While the diagram ↑ above right shows only a few dozen complexes, each disk actually contains thousands of such light detecting opsin complexes.)
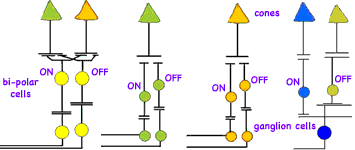
At the first synapses, bipolar cells have several types of receptors for the neurotransmitter glutamate, allowing these cells to respond to photoreceptor signals several ways. Some bipolar cells are tuned to faster and some to slower fluctuations in the visual signal; some glutamate receptors resensitize rapidly and others more gradually. The cells thus fire either quickly in succession or relatively slowly in response to the same amount of stimulation. These receptors respond to glutamate by activating what's known as an OFF pathway in the visual process, detecting dark images against a lighter background. (Recall that photoreceptors constantly release glutamate unless exposed to light.) Other bipolar cells have inhibitory glutamate receptors which prevent the bipolar cell from firing when the cell is exposed to the neurotransmitter. These receptors activate the ON pathway, detecting light images against a darker background. Our eyes have parallel visual circuits for ON (detecting light areas on dark backgrounds) and OFF (detecting dark areas on light backgrounds). Our vision (and that of other vertebrates) detects the contrast between images and their backgrounds. For example, we read black letters against a white background using the OFF circuits. Parallel ON/OFF bipolar cells transmit inputs to ganglion cells. This mechanism while it may seem complex at first, allows the eyes to distinguish contrasting objects over a much wider range of light conditions than otherwise possible.
Horizontal cells also receive input from cones at the first synaptic level in the retina. Each horizontal cell receives input from many cones, so its visual area is large. Their membranes also fuse with those of neighboring horizontal cells at gap junctions. As a result the electrical potentials of their membranes are the same over a very large area. Meanwhile, a single bipolar cell receives input from a handful of cones and thus has a medium-size receptive field. Whereas a single bipolar cell with its OFF or ON light response would carry a fairly blurry edge to its ganglion cell, horizontal cells add an opponent signal that is constricts the area of the bipolar cell's signal. Thus horizontal cells define precise edges to images and allows fine visual details. Horizontal cells also respond to signals from the inner plexiform layer transmitted via substances such as dopamine, nitric oxide and retinoic acid. The result is that horizontal cells adjust for different lighting conditions allowing signaling to become less sensitive in bright light and more sensitive in dim light.
The physiological properties of only a handful of retinal ganglion cell types have been studied in detail. One rare type of Y-cell are called upsilon cells. There is only about one upsilon cell for every 104 rods and cones. Each upsilon cell receives signals from approximately 100 light-sensing cells in its vicinity and combining these signals to generate an overall response for the region. But unlike most other feature-recognition cells which respond only to the overall brightness in their regions, the upsilon cells respond each time a pattern in their region changed, giving upsilon cells the ability to detect motion of viewed objects.
The ganglion cells have a receptive field organized as concentric circles. The amacrine cell circuitry in the inner plexiform layer conveys additional information to the ganglion cell, possibly sharpening the boundary between center and surround even further. In human retinas, two basic types of ganglion cells ON center and OFF center provide the major output of the retina. ON-center ganglion cells are activated when a spot of light falls in the center of their receptive field and are inactivated when light falls on the field's periphery. OFF-center ganglion cells react in the opposite way: Their activity increases when the periphery of their receptive field is lit and decreases when light falls on the center of the field.
In contrast to the rest of the retina, the center of vision contains midget ganglion cells connected in a one-to-one ratio with midget bipolar cells so carry information from a single cone to the brain. Each red or green cone in the central fovea connects to two midget ganglion cells, so at all times each cone can either transmit a dark-on-light (OFF) signal or a light-on-dark (ON) message. As a result, signals to the brain from this central area of vision carries both spatial and color information of the finest resolution. Messages from blue cones are not processed in the same way as from red and green cones, perhaps because the blue system evolved earlier. Blue cones transmit information through a special blue cone bipolar cell to a different type of ganglion cell, which can carry both a blue ON and a yellow OFF response.
The rod circuitry providing much of peripheral vision involves amacrine cells which sometimes stretch across the inner plexiform layer for hundreds of microns and interact with hundreds of bipolar cells and many ganglion cells collecting and amplifying very dim light for twilight and night vision. Usually, they emit the neurotransmitter glycine or GABA (gamma-aminobutyric acid). Most also release at least one other peptide such as substance P, somatostatin, vasointestinal peptide and cholecystokinin or neurotransmitter serotonin, dopamine, acetylcholine, adenosine and nitric oxide. Presumably these adjust for changing light intensities or have other effects. Intermittent flashing light stimulates a specialized amacrine cell to release dopamine uncoupling gap junctions between horizontal cells, reducing the size of their receptive fields. Dopamine also affects glutamate receptors on horizontal cells so that the amplitude of the light response declines. A type of ganglion cells which contain light reactive molecules melanopsins seem to function as their own giant photoreceptors, without needing input from rods or cones.
The common notion that human eyes have three color cones that feed primary color intensities to the brain (red, green, blue similar to that sent to a monitor or TV screen) is clearly inadequate. While there are only identical light detecting retinal molecules, small variations in the surrounding large opsin protein modify the sensitivity so that three variations in cones have overlying but slightly different band widths. But after processing, what generally is sent into the brain is comparisons between cone output. What is known as Opponent Color Theory suggests that perception of color is produced by several different comparisons between neighboring cones. Certain colors are never perceived together, i.e. they do not mix. We never see bluish-yellows or reddish-greens. This is consistent with the idea that color perception is based on comparisons between photoreceptor signals. For example, the yellow detector is always inactive when the blue detector is active and vice versa. A similar situation occurs for the neurons responding to red or green. The comparisons may be somewhat like the following:
negativeof the image and its color.
How can this be? Perhaps I am not far removed from Einstein's curved universe.
The May 1959 issue of Scientific American had an article on the color vision experiments by Edwin (Din
) Herbert Land (1909 - 1991), inventor of plastic sheets of Polaroid filters (such as in sun glasses), later the creator of the Polaroid instant camera and film, still later instant color photography, and the range finder system of using Doppler sound (used for self-opening store doors). The article suggested that our common belief that three primary colors are needed for full color perception is flawed!
 In 1955 Land (←at left) began studying the basics of color perception by repeating experiments first done by James Clerk Maxwell using 3 colors to form a full color image. But Land did the experiments with modern films and materials. On one occasion as he was dismantling his system of three projectors, he had turned off the blue-filtered projector, and removed the green filter from its projector. That left an image recorded through a red filter on the screen, projected with red light, and the green-recorded image, projected with white light. His assistant asked why all the colors in the original image were still visible! The image should have contained shades of black, white and red, but his assistant could see blue, green and yellow as well. After first glibly dismissing it, upon reflection Land returned to the laboratory and investigated further. When he darkened the image from the unfiltered projector, the colors became richer. He found that people often described colors of objects with no relationship to the intensities of each color actual reflected from those objects. His experiments helped lead to the understanding of color perception described above.
In 1955 Land (←at left) began studying the basics of color perception by repeating experiments first done by James Clerk Maxwell using 3 colors to form a full color image. But Land did the experiments with modern films and materials. On one occasion as he was dismantling his system of three projectors, he had turned off the blue-filtered projector, and removed the green filter from its projector. That left an image recorded through a red filter on the screen, projected with red light, and the green-recorded image, projected with white light. His assistant asked why all the colors in the original image were still visible! The image should have contained shades of black, white and red, but his assistant could see blue, green and yellow as well. After first glibly dismissing it, upon reflection Land returned to the laboratory and investigated further. When he darkened the image from the unfiltered projector, the colors became richer. He found that people often described colors of objects with no relationship to the intensities of each color actual reflected from those objects. His experiments helped lead to the understanding of color perception described above.
Read about and perhaps try reproducing Land's Experiments in Color Vision (web site by Wendy Carlos).
The Rowland institute at Harvard founded and endowed by Land, and where he continue scientific research, has additional information about Land. In 1963 he told MIT students, Science, to put it somewhat vulgarly, is a technique to keep yourself from kidding yourself. The only safe procedure for you, now that you have started, is to make sure that from this day forward until the day you are buried, you do two things each day. First, master a difficult old insight, and, second, add some new piece of knowledge to the world each day. Now does that seem extravagant?
To Land science isn't just a matter of trying things at random, but trying what is most likely to work first.
Communicating technical information such as observations and findings is a skill used by scientists but useful for most others. If you need course credit, use your observations in your journal to construct a formal report.
![]()
to next investigation
to Biochemistry menu
to ie-Chemistry menu
to site menu
{kind=link}